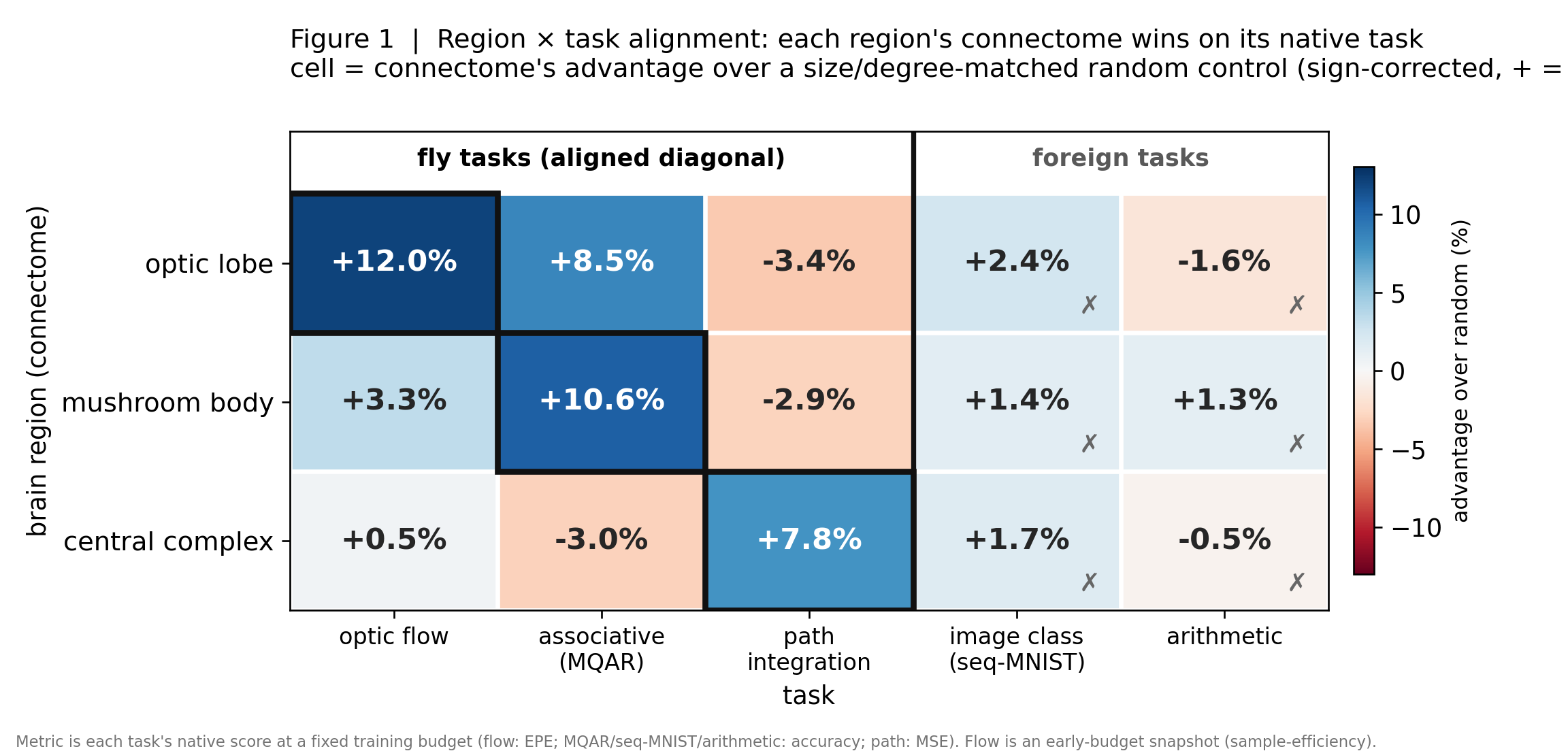
Real wiring beats random - but only at the job it evolved for
Three brain regions × five tasks. Each cell is the connectome's advantage over a matched-random control. Watch the diagonal light up - each region wins on the one task its circuit evolved to do - while the off-diagonal stays dark. That is a double dissociation.
Hover or tap any cell to read what it means. The gold-ringed cells are each region's native task.
The comparison is deliberately brutal: every real connectome is pitted against random networks matched on size, density and degree distribution, so the ~8-12% edge along the diagonal can only come from the specific pattern of connections. A diagonal that lights up while off-diagonal cells stay flat is the cleanest evidence that wiring carries task-specific computation rather than a generic boost.